

Fledging changes everything.
Wing clipping before fledging–the important developmental stage when a bird learns to fly–has the potential to inflict the most serious and long-lasting consequences compared with a singular clip of equal duration at any other time in its life. Wing clipping that interferes with fledging effectively interferes with the normal development of a young animal. Every bird owner, current or future, deserves to know how a bird can be affected by this.
If you visit a place which sells birds–depending upon which one you visit, of course–you may see baby birds in small pens, bins, cabinets, or cages. These enclosures are appropriate for baby birds who are not yet ready to leave the nest. The trouble arises, however, when baby birds are clipped and/or remain confined within small spaces past the age they should be learning to fly. In the wild, these fledgling parrots would be building flight skills, exploring their environments, following their parents, and learning how to be birds. To put it simply, the baby birds who are clipped and/or confined do not experience a normal transition to adulthood. I am going to show you why this is a very problematic and even unethical way of raising baby birds which is falling out of favour among reputable aviculturists. I understand that the people still following the outdated “wing clipping method” of rearing may, in many cases, be true bird-lovers who believe they are doing the right thing (or at least causing minimal harm), but that does not mean we need to accept or support it. There are many sources from which one can purchase parrots which have been raised flighted, but one must do one’s research. Private breeders, pet shops, and bird stores should all be taken as individual entities as their views on flight and wing clipping can vary tremendously.

At first glance, these enclosures may seem satisfactory. They’re clean. The birds have food, water, and even a few toys. But these birds, at an age when they should be flying and experiencing as much physical and mental stimulation as possible, are confined to tiny cubicles. What calibre of brain development can be derived from shuffling back and forth on a single perch? Credit: Anonymous contribution.
First, how does wing clipping before fledging affect the physical aspects of a bird’s development? A baby bird instinctively learns to walk, flap, and fly, similar to the way a baby human instinctively learns to crawl, “cruise”, and walk. The role of wings in avian locomotion is so deeply ingrained that baby birds adopt the characteristic flapping stroke associated with flight well before their musculoskeletal systems have finished developing. By using their legs and wings cooperatively while their muscles and bones develop, they are able to make a seamless transition to wing-based movement, or flight.1 Conversely, human babies use their arms and legs cooperatively to help them transition to leg-based movement.2 In short, a parrot’s wings do for them what our legs do for us.
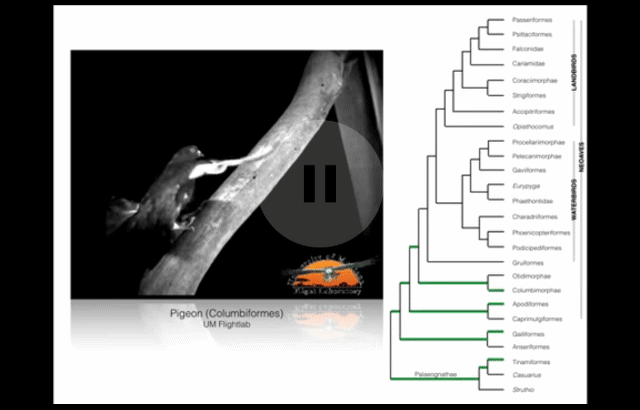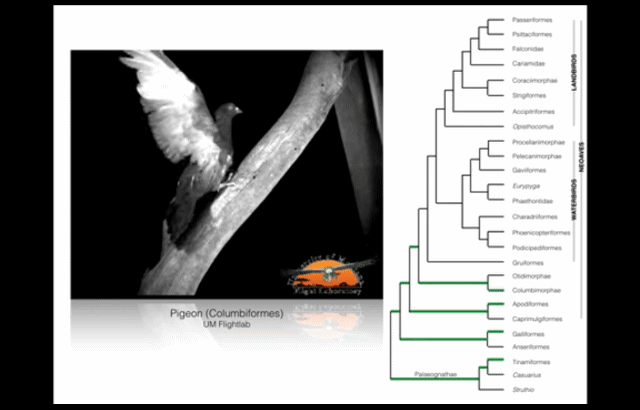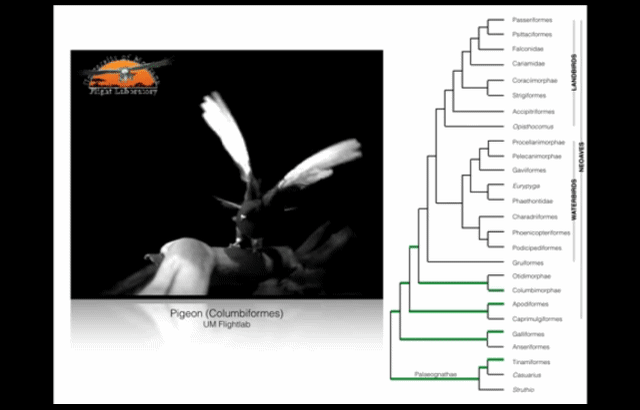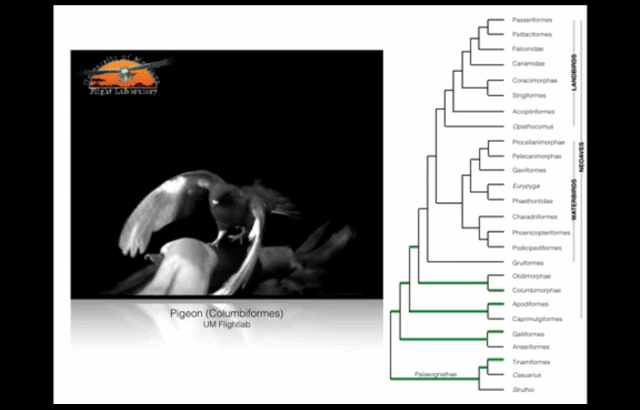
Wing-assisted incline running (WAIR) is a transitional behaviour that allows baby birds to supplement their underdeveloped wings and flight muscles with their legs. (Source: see footnote 1.) This video presents examples of WAIR throughout the avian “family tree”. Credit: UM Flight Lab on Youtube.
Without primary/flight feathers, however, the baby bird is unable to make this transition. Flight feathers strongly affect wingspan, reduce drag, and contribute greatly to the production of thrust and lift.3, 4 Bird wings do come in slightly different shapes, but certain characteristics, like primary feathers and wing tip shape, remain quite similar across avian species. These are “central to flight performance…because of their aerodynamic importance”.5 Thus, wing clipping and fundamentally altering their aerodynamic shape infringes upon a baby bird’s ability to learn normal locomotion. How can a baby bird, whose flight muscles are still developing, achieve flight without a properly shaped wing? How can it adequately build its flight muscles? A recent study on pigeons found that the pectoralis muscle grew rapidly after fledging, all the way up until sexual maturity.6

This clipped baby cockatiel is given the opportunity to attempt flight but, due to the sheer aerodynamic difficulty coupled with weak flight muscles, drops straight to the floor. Credit: @peace_of_parrots on Instagram.
If muscles aren’t properly developed and remain weak, they can’t place an appropriate burden on the bones they control, leading to negative effects on bone mass, size, and strength.7 The mechanical load placed on a bone while it is growing ultimately determines the strength of that bone8, 9, and the mechanical load imposed by wing exercise at the onset of flight prompts specific and rapid changes in bone strength.10 It stands to reason that if a bird’s wing bones do not experience the burden of flight during development, they will be weaker than those which have been subjected to flight during development. Additionally, exercise during growth results in bones of higher mass and density.11, 12 If exercise continues throughout the animal’s life, this increased initial bone strength can help reduce the risk of fractures as it ages.13

While a bird may reasonably stay in a cage of appropriate size for part of the day and then come out for flighted exercise, most “bird store” birds for sale do not leave their cages. If they do, their wings are clipped. This is detrimental to both their development and their relationship with humans. Birds of fledging age and onwards, even ones “for sale”, should be housed in cages or aviaries that allow for flighted exercise. Alternatively, they should receive daily flighted exercise outside their cages. Birds should have a variety of perches (not dowels), and should furthermore have adequate enrichment available in the form of toys (e.g. foraging, shredding, interactive) and/or natural, bird-safe foliage and branches. If we want our birds to receive the best upbringing possible, we must be critical of the environment in which they are raised. We should ask ourselves how birds can be provided developmental opportunities which come closer to those they would experience in nature, even if we can’t replicate them perfectly. Credit: Anonymous contribution.
The effects of wing clipping on physical development may seem troubling, but perhaps even more troubling are the effects of wing clipping on brain development. During an animal’s early life, there exist sensitive periods wherein its experiences (or lack thereof) have a profound effect on its brain.14 Animals raised in enriched environments which incorporate mental, physical, and social stimulation have more active, plastic brains, possessing a greater number of, and more complex, neural connections.15, 16 The baby bird navigating its environment by flying is learning a series of complex motor skills: taking off, flapping, gliding, ascending, descending, turning, avoiding obstacles, braking, landing, and so on. In doing so, it is quickly interpreting visual data. Compare this baby bird, who is following a normal developmental path, to the baby bird in the bin who cannot move beyond the same stationary flapping it would have done while still in the nest. Studies point to a significant relationship between physical activity, motor skills, and overall cognitive function.17 Research also indicates that the presence or absence of physical exercise can determine just how enriched an environment is.

This fledgling macaw is learning indoor flight and recall. In doing so, he is building muscle, establishing neural pathways, and forming a positive association with humans. Accomplishing this flight-based task would be impossible with clipped wings. Credit: @flightedfeathers on Instagram.
To quote a recent study conducted on mice, which are the standard lab animals for experiments in neuroscience, “As the brain matures, its ability to change declines, together with the ability to learn, memorize or recover from various brain injuries. One of the approaches shown to improve brain plasticity is enriched environment (EE)”.18 The study compared mice raised in standard cages to those raised in an enriched environment, one which was nine times larger than the standard cage, had two floors, a ladder, a maze, a slide, a tunnel, and three running wheels. Scientists discovered that even a few days of being housed in a standard cage without a running wheel completely eliminated the type of brain plasticity they were studying, while the addition of a running wheel was able to preserve it. The mice in the standard cages were able to walk but not run, similar to a clipped baby bird who is able to walk but not fly. Biologically appropriate, sufficiently demanding physical exercise made the difference. Studies also show that physical activity during critical periods results in better learning, concentration, memory, and intelligence. (Source: see footnote 17.) Based on this information, is it not logical to conclude that the baby bird who learns to fly will receive higher levels of environmental enrichment and exercise, and will therefore achieve a higher level of brain development, than the baby bird who is flight-restricted?

No enriching physical exercise is possible for the birds in these cages. They are clipped, never learned how to fly, do not leave their cages, or all of the above. Displays like this teach people to see perch-bound parrots as “normal”. Changes which work towards accommodating flight can and should be made by the owners of these establishments. Credit: Anonymous contribution.
It is impossible to know for sure whether a particular bird’s behavioural issues are due to this type of rearing or another cause entirely, but we can make the logical inference that abnormal development increases the incidence of abnormal behaviour. Clipped baby birds are often advertised as “hand-raised” or “tame”, but many of them are fearful and averse to human contact. The shortcut which circumvents socialising and training these birds is to physically disable them via wing clipping. Because they are unable to fly, they cannot escape a customer who wants to hold them regardless of how fearful they may be. When a bird is afraid, repeatedly forcing it to interact with us and removing all avenues of avoidance does not result in tameness or “bonding”; it results in learned helplessness. Learned helplessness is a condition where an animal or person is first confronted by inescapable stress or pain. Then, later on, even if it would be possible for them to escape the stress or pain, they don’t make any attempt to.19 Essentially, if we force a clipped bird to step up over and over, or if we pick it up off the ground every time it tries to flutter away, we are forcing it into a state of learned helplessness. It’s still afraid, but it has given up. This naturally leads to a more insecure, dependent bird who feels safe in a very limited range of situations–a “one-person bird” in the making.

Baby birds are incredibly easy to train. Giving them the choice to interact with us in exchange for rewards, as well as the freedom to decline interaction, fosters a relationship based on cooperation rather than force. Credit: @oakeyandcrew on Instagram.
The other, equally unfortunate, result of forcing interactions with birds is that some of them learn that when flying away is futile, biting works very well. Instead of turning to their instinctive avoidance strategy, flight, they rely on other behaviours, like biting, to try and make perceived threats go away. Flight-restricted birds can’t build the confidence that comes of having the power to fly away from something they don’t like. Fully flighted birds can and do bite in many contexts, but they do not have the added stressor of a physical handicap or the psychological insecurity caused by that handicap encouraging them to bite in instances where they would otherwise elect not to. Dr. Michael Doolen, avian veterinarian, suspects that a great deal of problem behavior he has seen in his patients results from “imprinted birds not being allowed to fledge and develop strong flight skills”.20 He has also observed that flight skills cultivated later in life can reduce or resolve some of this problem behaviour.

Interactions of this kind with eager customers teach some flight-restricted baby birds that biting hard enough will drive hands away. The bird is clearly uncomfortable, but customers–many of whom lack knowledge of bird behaviour–are led to believe that these birds are hand-tame and friendly. Wing clipping is still overwhelmingly common with many bird stores and breeders. Credit: Anonymous contribution.
The rearing of baby birds is obviously a complex issue and many factors, not just wing clipping, are at work here. However, the denial of the fledging process most certainly influences a baby bird’s development by barring it from normal exercise and stimulation. You may now wonder: can birds learn to fly if they were clipped before fledging? After all, the adult brain does maintain a certain degree of plasticity. (Source: see footnote 15). Fortunately, they can usually learn to fly, and there are many people all over the world helping their clipped or previously clipped birds do just that. But it isn’t as easy for these birds, and even with a great deal of effort, they may never become as skilled as birds who fledged at the natural age.

Birds who have never learned how to fly, or who have been kept flight-restricted, must be gradually taught and/or rehabilitated. Notice how this Hahn’s macaw leans in the direction he wants to go but is mentally unable to bring himself to fly. It was necessary to teach him how to first step, then hop, then hop and flap onto a perch before he could progress to true flight. This process requires patience and dedication on the part of the bird’s owner. Some birds learn faster than others, of course; larger and heavy-bodied parrots tend to have the most difficulty. This Hahn’s macaw has achieved flight with the help of his new owner, but he is not yet a strong or coordinated flyer. Credit: @flightedfeathers on Instagram.
Why might this be? If flight is natural, why is it so hard for these birds to learn later in life? Well, there are a few reasons. During sensitive periods of brain development, our experiences help shape our abilities. The developing brain forms an overabundance of connections and is constantly making new ones based on the information it receives. The connections being used become stronger, while the ones left unused are eliminated.21 Genie, a “feral child” who was not exposed to language from the age of about two to thirteen, ultimately did learn some spoken language but was never able to harness the grammatical skills normal children learn by the age of five.22 Another individual, who was merely deaf and uneducated up until the age of fifteen (rather than abusively isolated as in Genie’s case), showed striking similarities to Genie in his language deficits. After almost three years, he was still unable to comprehend verb tense, negation, pronouns, or prepositions.23 People who learn to play musical instruments as young children acquire structural differences in their brains compared to non-musicians. These differences allow them to perform better at certain motor tasks, musical tasks, and even language processing tasks.24 Rats raised in the reduced gravity of low Earth orbit have more difficulty maintaining balance and righting themselves when they return to Earth compared to rats raised in normal gravity.25 All of these examples, though of course not perfectly analogous with fledging, illustrate long-lasting or permanent effects associated with sensitive periods of brain development.

This 1.5-year-old macaw was clipped before fledging and previously confined to a small cage. It took almost five months of gradually building her strength and confidence before she achieved her first short-distance recall. Her journey to becoming a trained free-flyer has proven far more arduous than that of her owners’ other macaw, who fledged normally and was never clipped. Credit: Mikey the Macaw on Youtube; @mikey_themacaw on Instagram.
The ability to learn and acquire new skills obviously continues throughout life, as evidenced by people who learn to play instruments as adults and by birds who learn to fly as adults. However, the developmental periods before puberty are windows of opportunity that are not to be underestimated. One paper on motor learning in children concludes that these time periods should “be used particularly intensively…especially with regard to coordination and speed”.26 Although we don’t have any studies on how flight restriction during sensitive periods may impact flight later in life, we do have studies on other kinds of sensitive periods in birds, such as sexual imprinting,27 sensorimotor integration during song-learning,28 and auditory-spatial learning,29 as proof of concept. Because a clipped baby bird is unable to fly, its brain is unable to build the specific connections that flight would have “unlocked”. Building these connections later, when the brain is no longer at peak plasticity, can pose a serious challenge. Thus, wing clipping significantly complicates a process–learning to fly skillfully–that would otherwise be natural and intuitive.
This is especially true when it comes to outdoor free flight. Many birds (particularly larger birds like macaws and cockatoos) have been trained, with professional help, for outdoor free flight even if they were clipped before fledging. I think it is wonderful that those birds and trainers have accomplished such a feat. However, the risk and difficulty level when training these birds outdoors increases substantially compared to training a bird who has experienced normal development; such an undertaking is certainly not ideal. As free flight becomes more common and more people wish to fly parrots, it is paramount that the importance of fledging and the effects of wing clipping (in addition to working with a highly experienced mentor) be understood.
How can wing clipping affect physical health throughout a bird’s life?
References
- Heers, A.M., Baier, D.B., Jackson, B.E. and Dial, K.P., 2016. Flapping before flight: high resolution, three-dimensional skeletal kinematics of wings and legs during avian development. PLoS One, 11(4), p.e0153446. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0153446
- Adolph, K.E., Berger, S.E. and Leo, A.J., 2011. Developmental continuity? Crawling, cruising, and walking. Developmental science, 14(2), pp.306-318. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3050596/
- Shatkovska, O.V. and Ghazali, M., 2017. Relationship between developmental modes, flight styles, and wing morphology in birds. The European Zoological Journal, 84(1), pp.390-401. https://www.tandfonline.com/doi/full/10.1080/24750263.2017.1346151
- Dvořák, R., 2016. Aerodynamics of bird flight. In EPJ Web of Conferences (Vol. 114, p. 01001). EDP Sciences. https://www.epj-conferences.org/articles/epjconf/pdf/2016/09/epjconf_efm2016_01001.pdf
- Wang, X. and Clarke, J.A., 2015. The evolution of avian wing shape and previously unrecognized trends in covert feathering. Proceedings of the Royal Society B: Biological Sciences, 282(1816), p.20151935. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4614784/
- Liang, X., Yu, J., Wang, H. and Zhang, Z., 2018. Post‐Hatching Growth of the Pectoralis Muscle in Pigeon and Its Functional Implications. The Anatomical Record, 301(9), pp.1564-1569. https://www.ncbi.nlm.nih.gov/pubmed/29729220
- Going, S.B. and Farr, J.N., 2010. Exercise and bone macro-architecture: is childhood a window of opportunity for osteoporosis prevention?. International journal of body composition research, 8, p.1. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3903297/
- Bergmann, P., Body, J.J., Boonen, S., Boutsen, Y., Devogelaer, J.P., Goemaere, S., Kaufman, J., Reginster, J.Y. and Rozenberg, S., 2010. Loading and skeletal development and maintenance. Journal of osteoporosis, 2011. https://www.hindawi.com/journals/jos/2011/786752/
- Regmi, P., Deland, T.S., Steibel, J.P., Robison, C.I., Haut, R.C., Orth, M.W. and Karcher, D.M., 2015. Effect of rearing environment on bone growth of pullets. Poultry science, 94(3), pp.502-511. https://academic.oup.com/ps/article/94/3/502/1520001
- Habib, M.B. and Ruff, C.B., 2008. The effects of locomotion on the structural characteristics of avian limb bones. Zoological Journal of the Linnean Society, 153(3), pp.601-624. https://academic.oup.com/zoolinnean/article/153/3/601/2606425
- Karlsson, M., Nordqvist, A. and Karlsson, C., 2008. Physical activity increases bone mass during growth. Food & nutrition research, 52(1), p.1871. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2596740/
- Biewener, A.A. and Bertram, J.E., 1994. Structural response of growing bone to exercise and disuse. Journal of Applied Physiology, 76(2), pp.946-955. https://www.physiology.org/doi/abs/10.1152/jappl.1994.76.2.946
- Colì, G., 2013. To prevent the osteoporosis playing in advance. Clinical Cases in Mineral and Bone Metabolism, 10(2), p.83. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3797006/
- Dulac, O., Lassonde, M., Sarnat, H.B. (eds.), 2013. Pediatric Neurology, Part 1. Newnes, p. 4. https://books.google.com/books?id=IWnXvarDNywC&dq
- Bengoetxea, H., Ortuzar, N., Bulnes, S., Rico-Barrio, I., Lafuente, J.V. and Argandona, E.G., 2012. Enriched and deprived sensory experience induces structural changes and rewires connectivity during the postnatal development of the brain. Neural plasticity, 2012. https://www.hindawi.com/journals/np/2012/305693/
- Markham, J.A. and Greenough, W.T., 2004. Experience-driven brain plasticity: beyond the synapse. Neuron glia biology, 1(4), pp.351-363. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1550735/
- Zeng, N., Ayyub, M., Sun, H., Wen, X., Xiang, P. and Gao, Z., 2017. Effects of physical activity on motor skills and cognitive development in early childhood: a systematic review. BioMed research international, 2017. https://www.hindawi.com/journals/bmri/2017/2760716/
- Kalogeraki, E., Pielecka-Fortuna, J. and Löwel, S., 2017. Environmental enrichment accelerates ocular dominance plasticity in mouse visual cortex whereas transfer to standard cages resulted in a rapid loss of increased plasticity. PloS one, 12(10), p.e0186999. https://journals.plos.org/plosone/article?id=10.1371/journal.pone.0186999
- Song, L., Che, W., Min-Wei, W., Murakami, Y. and Matsumoto, K., 2006. Impairment of the spatial learning and memory induced by learned helplessness and chronic mild stress. Pharmacology Biochemistry and Behavior, 83(2), pp.186-193. https://www.sciencedirect.com/science/article/pii/S0091305706000086
- Velasco, M.C., 2012. Are there long-term effects of production-based rearing on pet bird behavior?. Veterinary Clinics: Exotic Animal Practice, 15(2), pp.205-214. https://www.vetexotic.theclinics.com/article/S1094-9194(12)00014-X/fulltext
- Tierney, A.L. and Nelson III, C.A., 2009. Brain development and the role of experience in the early years. Zero to three, 30(2), p.9. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3722610/
- Pines, M., 1997. The civilizing of Genie. Teaching English through the Disciplines: Psychology. New York, NY: Whittier. Available at: http://www.geocities.ws/drobste/genie.doc
- Grimshaw, G.M., Adelstein, A., Bryden, M.P. and MacKinnon, G.E., 1998. First-language acquisition in adolescence: Evidence for a critical period for verbal language development. Brain and Language, 63(2), pp.237-255. https://canlabvuw.files.wordpress.com/2015/05/first-language-acquisition-in-adolescence.pdf
- White, E.J., Hutka, S.A., Williams, L.J. and Moreno, S., 2013. Learning, neural plasticity and sensitive periods: implications for language acquisition, music training and transfer across the lifespan. Frontiers in systems neuroscience, 7, p.90. https://www.frontiersin.org/articles/10.3389/fnsys.2013.00090/full
- Harding, S.M., Singh, N.J. and Walton, K.D., 2017. A Sensitive Period for the Development of Motor Function in Rats: A Microgravity Study. Gravitational and Space Research, 5(2). http://gravitationalandspacebiology.org/index.php/journal/article/view/767/795
- Hirtz, P. and Starosta, W., 2002. Sensitive and critical periods of motor co-ordination development and its relation to motor learning. Journal of human kinetics, 7, pp.19-28. http://www.johk.pl/files/03_hir.pdf
- Crawford, W.C., The Theory of Imprinting-Its Implications and Ramifications in Raptors and All Birds. AFA Watchbird, 18(2), pp.34-35. https://journals.tdl.org/watchbird/index.php/watchbird/article/view/2638
- Pytte, C.L. and Suthers, R.A., 2000. Sensitive period for sensorimotor integration during vocal motor learning. Journal of neurobiology, 42(2), pp.172-189. https://www.ncbi.nlm.nih.gov/pubmed/10640325
- Knudsen, E.I., 2002. Instructed learning in the auditory localization pathway of the barn owl. Nature, 417(6886), p.322. https://web.mit.edu/kardar/www/research/seminars/CorticalMaps/DynaPage.pdf